
Shark. Sharks are a group of elasmobranch fish characterized by a cartilaginous skeleton, five to seven gill slits on the sides of the head, and pectoral fins that are not fused to the head.
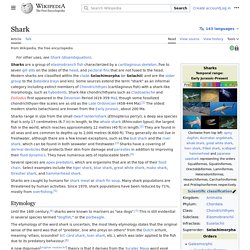
Modern sharks are classified within the clade Selachimorpha (or Selachii) and are the sister group to the rays. However, the term "shark" has also been used for extinct members of the subclass Elasmobranchii outside the Selachimorpha, such as Cladoselache and Xenacanthus, as well as other Chondrichthyes such as the holocephalid eugenedontidans. Under this broader definition, the earliest known sharks date back to more than 420 million years ago.[1] Acanthodians are often referred to as "spiny sharks"; though they are not part of Chondrichthyes proper, they are a paraphyletic assemblage leading to cartilaginous fish as a whole.
Since then, sharks have diversified into over 500 species. Etymology Evolution Most fossil sharks from about 300 to 150 million years ago can be assigned to one of two groups. Tetrapod. While most species today are terrestrial, little evidence supports the idea that any of the earliest tetrapods could move about on land, as their limbs could not have held their midsections off the ground and the known trackways do not indicate they dragged their bellies around.
Presumably, the tracks were made by animals walking along the bottoms of shallow bodies of water.[2] Amphibians today generally remain semiaquatic, living the first stage of their lives as fish-like tadpoles. Several groups of tetrapods, such as the snakes and cetaceans, have lost some or all of their limbs. In addition, many tetrapods have returned to partially aquatic or fully aquatic lives throughout the history of the group (modern examples of fully aquatic tetrapods include cetaceans and sirenians). A quadruped actually uses four limbs for locomotion; not all tetrapods are quadrupeds. Biodiversity[edit] Tetrapoda includes four classes: amphibians, reptiles, mammals, and birds.
Evolution[edit] Placodermi. The first identifiable placoderms appear in the fossil record during the late Llandovery epoch of the early Silurian.[4] The various groups of placoderms were diverse and abundant during the Devonian, but became extinct at the end-Devonian Hangenberg event 360 million years ago [5] Characteristics[edit] One of the largest known arthrodires, Dunkleosteus telleri, was 8–11 m (26–36 ft) long, and is presumed to have had a large distribution, as its remains have been found in Europe, and North America and possibly Morocco.
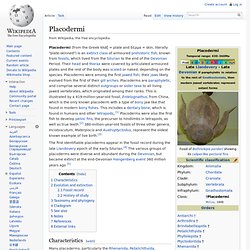
Some paleontologists regard it as the world's first vertebrate "superpredator", preying upon other predators. Osteichthyes. Characteristics[edit] Bony fish have no placoid scales.
Mucus glands coat the body. Most have smooth and overlapping scales of that are ganoid, cycloid or ctenoid. Classification[edit] This has led to an alternative classification, splitting the Osteichthyes into two full classes. Sarcopterygii. Characteristics[edit] Guiyu oneiros, the earliest known bony fish, lived during the Late Silurian, 419 million years ago).[1][2] It has the combination of both ray-finned and lobe-finned features, although analysis of the totality of its features place it closer to lobe-finned fish.[3][4][5][6] Most species of lobe-finned fishes are extinct.
The largest known lobe-finned fish was Rhizodus hibberti from the Carboniferous period of Scotland which may have exceeded 7 meters in length. Among the two groups of extant (living) species, the coelacanths and the lungfishes, the largest species is the West Indian Ocean coelacanth, reaching 2 m (6.5 ft) in length and weighing up 110 kg (240 lb).
The largest lungfish is the African lungfish which can reach 2 m (6.6 ft) in length and weigh up to 50 kg (110 lb).[9][10] Actinopterygii. The ray-finned fishes are so called because they possess lepidotrichia or "fin rays", their fins being webs of skin supported by bony or horny spines ("rays"), as opposed to the fleshy, lobed fins that characterize the class Sarcopterygii which also, however, possess lepidotrichia.
These actinopterygian fin rays attach directly to the proximal or basal skeletal elements, the radials, which represent the link or connection between these fins and the internal skeleton (e.g., pelvic and pectoral girdles). In terms of numbers, actinopterygians are the dominant class of vertebrates, comprising nearly 99% of the over 30,000 species of fish.[1] They are ubiquitous throughout freshwater and marine environments from the deep sea to the highest mountain streams.
Extant species can range in size from Paedocypris, at 8 mm (0.3 in), to the massive ocean sunfish, at 2,300 kg (5,070 lb), and the long-bodied oarfish, at 11 m (36 ft). Characteristics[edit] Ray-finned fishes occur in many variant forms. Pteridospermatophyta. The term Pteridospermatophyta (or "seed ferns") refers to several distinct groups of extinct seed-bearing plants (spermatophytes).
Pteridophyte. Pteridophytes or Pteridophyta, in the broad interpretation of the term (or sensu lato), are vascular plants (plants with xylem and phloem) that reproduce and disperse via spores.
Because they produce neither flowers nor seeds, they are referred to as cryptogams. The group includes ferns, horsetails, clubmosses, spikemosses and quillworts. These do not form a monophyletic group, because ferns and horsetails are more closely related to seed plants than to lycophytes (clubmosses, spikemosses and quillworts). Therefore, pteridophytes are no longer considered to form a valid taxon, but the term is still used as an informal way to refer to ferns (monilophytes) and lycophytes, and some recent authors have used the term to refer strictly to the monilophytes.
Pteridophyte classification[edit] In addition to these living groups, several groups that are now extinct and known only from fossils are considered to belong to pteridophytes. Pteridophyte life cycle[edit] Proetida. Proetida is an order of trilobite that lived from the Ordovician to the Permian.
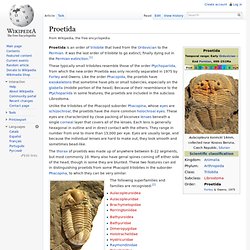
It was the last order of trilobite to go extinct, finally dying out in the Permian extinction.[1] These typically small trilobites resemble those of the order Ptychopariida, from which the new order Proetida was only recently separated in 1975 by Fortey and Owens. Like the order Phacopida, the proetids have exoskeletons that sometime have pits or small tubercles, especially on the glabella (middle portion of the head).
Because of their resemblance to the Ptychopariida in some features, the proetids are included in the subclass Librostoma. The thorax of proetids was made up of anywhere between 8–22 segments, but most commonly 10. Euramerica. Euramerica (also known as Laurussia (not to be confused with Laurasia), the Old Red Continent or the Old Red Sandstone Continent) was a minor supercontinent created in the Devonian as the result of a collision between the Laurentian, Baltica, and Avalonia cratons (Caledonian orogeny). 300 million years ago in the Late Carboniferous tropical rainforests lay over the equator of Euramerica.
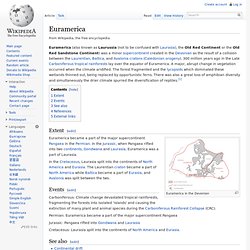
A major, abrupt change in vegetation occurred when the climate aridified. The forest fragmented and the lycopsids which dominated these wetlands thinned out, being replaced by opportunistic ferns. Late Devonian extinction. Millions of years ago Late D. Antler orogeny. Two facies of lower Paleozoic rocks[edit] Plate tectonics[edit] From an early date,[7] geologists have struggled to explain the presence in Nevada and adjacent areas of the Antler orogenic deposits without achieving a consensus. The advent of plate tectonic theory provided a variety of possible mechanisms by which the Roberts Mountains thrust and the orogenic deposits could be explained, but none of them has been universally accepted. Trilobite. Trilobites had many life styles; some moved over the sea-bed as predators, scavengers or filter feeders and some swam, feeding on plankton.
Most life styles expected of modern marine arthropods are seen in trilobites, with the possible exception of parasitism (where there are still scientific debates).[4] Some trilobites (particularly the family Olenidae) are even thought to have evolved a symbiotic relationship with sulfur-eating bacteria from which they derived food.[5] Fossil record[edit] All Olenellina lack facial sutures (see below), and this is thought to represent the original state. Ammonoidea. Ammonites are excellent index fossils, and it is often possible to link the rock layer in which a particular species or genus is found to specific geological time periods.
Their fossil shells usually take the form of planispirals, although there were some helically spiraled and nonspiraled forms (known as heteromorphs). Diagnostic characters[edit] Adaptive radiation. Four of the 14 finch species found on the Galápagos Archipelago, are thought to have evolved by an adaptive radiation that diversified their beak shapes to adapt them to different food sources. In evolutionary biology, adaptive radiation is a process in which organisms diversify rapidly into a multitude of new forms, particularly when a change in the environment makes new resources available, creates new challenges and opens environmental niches.[1][2] Starting with a recent single ancestor, this process results in the speciation and phenotypic adaptation of an array of species exhibiting different morphological and physiological traits with which they can exploit a range of divergent environments.[2] Identification[edit] Four features can be used to identify an adaptive radiation:[2] A common ancestry of component species: specifically a recent ancestry.
Devonian.